
У дійних корів, які страждають від теплового стресу, часто спостерігається зменшення жирності молока, що може завдавати збитків виробникам у складних умовах нинішнього молочного ринку. Проте поточне дослідження свідчить, що оптимізація функції рубця може сприяти підтримці концентрації молочного жиру в корів при тепловому стресі.
Синтез молочного жиру залежить від жирних кислот, які надходять з 2 джерел:
- Довголанцюгові жирні кислоти (більше 16 атомів вуглецю на молекулу) — виробляються від поглинання циркулюючих жирних кислот, харчових жирів, що всмоктуються з травного тракту, і нееcтерифікованих жирних кислот (НЕЖК) з мобілізованих запасів жиру.
- Коротколанцюгові (від 4 до 8 атомів вуглецю) і середньоланцюгові (від 10 до 14 атомів вуглецю) жирні кислоти — утворюються в молочній залозі за допомогою синтезу «de novo» (жирні кислоти, створюються «заново» в молочній залозі з менших молекулах).
Довголанцюгові жирні кислоти можуть походити з обох джерел. У добре нагодованої корови, за оцінками, від 4% до 8% молочних жирних кислот походять від розпаду жирових відкладень (наприклад, НЕЖК). Однак частка жирних кислот з цього джерела може поступово збільшуватися, оскільки зменшується чистий енергетичний баланс корови (Bauman and Griinari, 2001). Через дію теплового стресу існує два потенційних механізми зменшення молочного жиру. Перший — біогідрогенізація жирних кислот рубця — стримування новоутворення молочного жиру. Другий — ліпополісахариди рубця — обмеження постачання субстрату і новоутворення молочного жиру.
Змінена біогідрогенізація жирних кислот
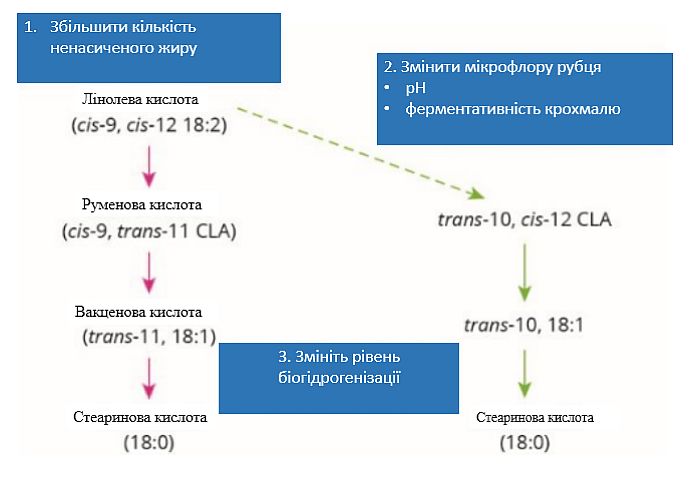
Згідно з загальноприйнятою «теорією біогідрогенізації» (Bauman and Griinari, 2001), зменшення молочного жиру виникає внаслідок змін у рубці біогідрогенізації ненасичених жирних кислот і проходження певних проміжних сполук біогідрогенізації з рубця (наприклад, транс-10, цис-12 CLA). Ці проміжні продукти біогідрогенізації згодом перешкоджають експресії генів, що беруть участь у синтезі жирів, тим самим зменшуючи синтез молочного жиру у вимені. Крім того, підвищена швидкість відтоку кормів з рубця може збільшити ймовірність проходження проміжних сполук біогідрогенізації через рубець. Таким чином, теорія визначає, яким чином деякі корми можуть бути факторами ризику зменшення молочного жиру (схема 1).
Нутриціоністи пропонують під час теплового стресу згодовувати додатковий жир для підтримки споживання енергії корів. Однак важливо, щоб жир був захищений від розпадання в рубці. В іншому випадку низький рН рубця, який утворюється у корів під дією теплового стресу, може генерувати більше проміжних сполук біогідрогенізації і збільшити ризик зменшення жирності молока.
Схема 1. Кормові добавки можуть впливати на ризик зменшення молочного жиру трьома шляхами через біогідрогенізацію рубця
(адаптовано з Lock та Bauman, 2007)
Змінене виробництво ліпополісахаридів рубця
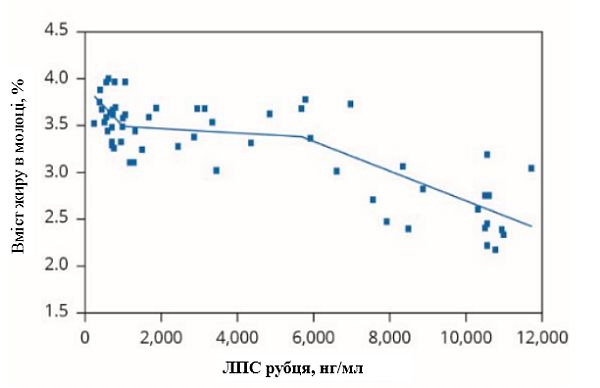
Інший потенційний механізм зменшення жиру в молоці під час теплового стресу включає концентрацію ліпополісахариду в рубці (ЛПС), який утворюється з грамнегативних бактерій, коли вони гинуть. Результати досліджень свідчать, що при зниженні рН рубця концентрація ЛПС в ньому підвищується. Відповідно, коли ЛПС підвищується, концентрація молочного жиру знижується. Zebeli та Ametaj (2009) показали, що концентрація ЛПС в рубці збільшується по мірі збільшення частки зерна в кормі. При збільшенні ЛПС вміст молочного жиру зменшується (графік 1). Цей взаємозв'язок може бути обумовлений здатністю ЛПС ініціювати вироблення інсуліну в підшлунковій залозі (Waldron et al., 2006). Підвищена циркуляція інсуліну і підвищена чутливість до нього під час теплового стресу може зменшити мобілізацію жиру. Ця умова може виникнути у корів, які страждають від теплового стресу, навіть попри негативний енергетичний баланс, спричинений зменшенням споживання корму та збільшенням вимог для життєзабезпечення (Baumgard and Rhoads, 2013). Крім того, недостатність НЕЖК в плазмі крові, потенційно важливого попередника синтезу молочного жиру під час теплового стресу (Bauman and Griinari, 2001), може сприяти зменшенню молочного жиру. Ще один негативний вплив ЛПС на виробництво жирних кислот включає: зниження активності ліпопротеїнової ліпази (Lopez-Soriano і Williamson, 1994), зменшення біосинтезу ліпопротеїнової ліпази та транспортного білка жирних кислот 1 (Feingold et al., 2009) і пригнічення ферментів, пов'язаних з новоутворенням жирних кислот в тканинах вимені (Dong et al., 2011).
Графік 1. Взаємозв'язок між ліпополісахаридом рубця (ЛПС) і вмістом жиру в молоці
(адаптовано з Zebeli та Ametaj, 2009)
Оптимізація функції рубця під час теплового стресу
Дане дослідження пояснює, як зменшення жирності молока під час теплового стресу залежить від здоров'я рубця. Враховуючи цей зв'язок, оптимізація функції рубця може допомогти зберегти вміст молочного жиру та ефективність виробництва молока у дійних корів під час теплового стресу.
Тепловий стрес спричинює фізіологічні та поведінкові зміни у дійних корів. Ці зміни можуть призвести до субоптимальних умов рубця, в результаті чого продукуються проміжні продукти біогідрогенізації жирних кислот і ЛПС, які стримують синтез молочного жиру у вимені.
На сьогодні доступні природні технології, які допомагають оптимізувати умови в рубці за допомогою раціону, тим самим сприяючи зменшенню негативного впливу теплового стресу на молочний жир і зберегти ефективність виробництва високопродуктивних молочних корів. Один із способів знизити падіння жиру в молоці при тепловому стресі -— введення в раціон захищених від розпаду в рубці жирів.
milkUa.Info за матеріалами dairyglobal.net